Studier i rhododendron
DNA -> stamtræ
Rhododendronslægtens kendetegn – Erica familien
Ordlister; dansk morphologi, engelsk taxonomi
STUDIER
Morphological studies in the genus Rhododendron. Dealing with fruits, seeds and seedlings and their associated hairs
ISBN 87-12-30853-6 vol I + II copyright by Johannes Hedegaard 1980.
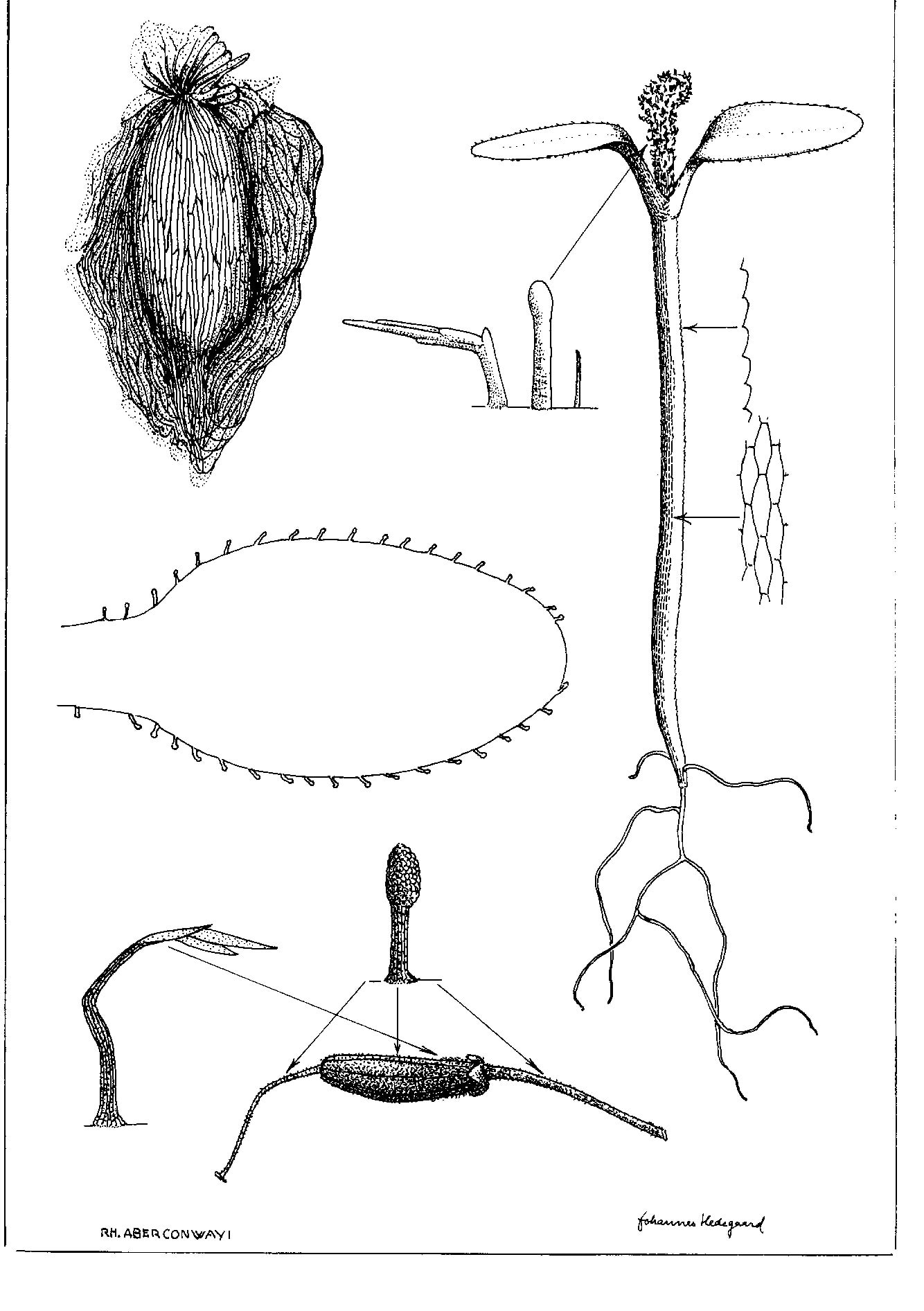
Drawings and text from about 300 different species. R. aberconwayi as an example.
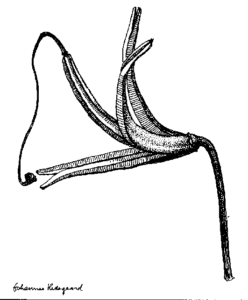
- R. aberconwayi: seed, fruit, hairs and seedling
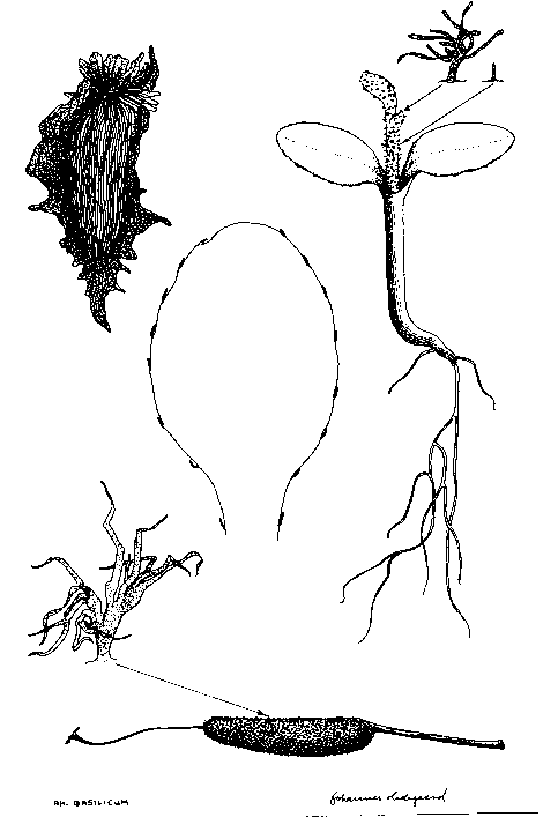
- R. basilicum: seed, fruit, hairs and seedling
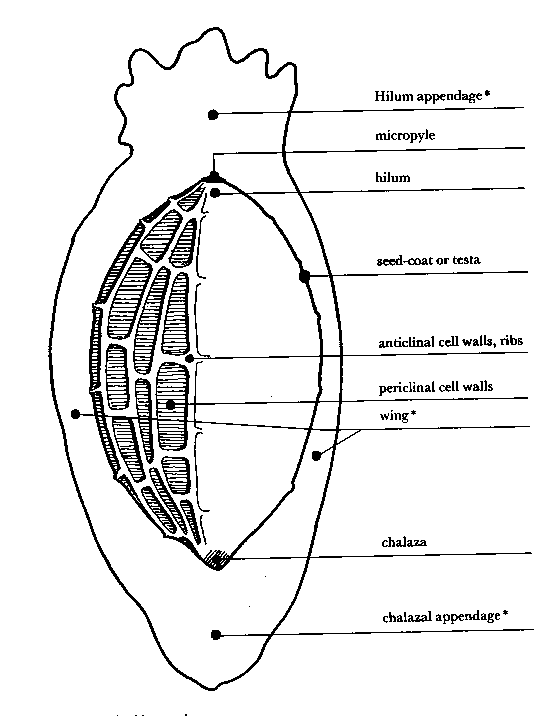
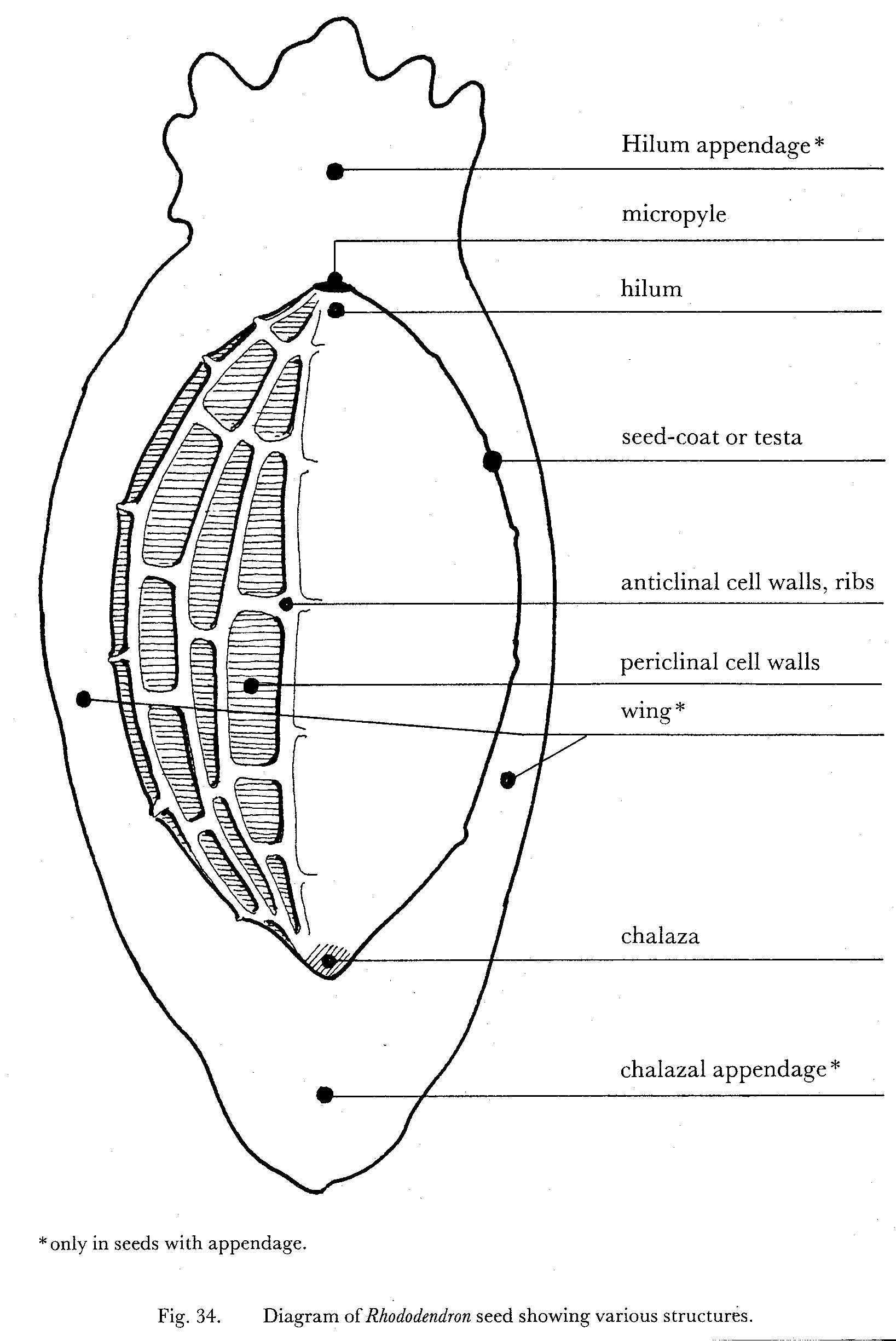
- Transverse section of a seed
- Different examples of calyx
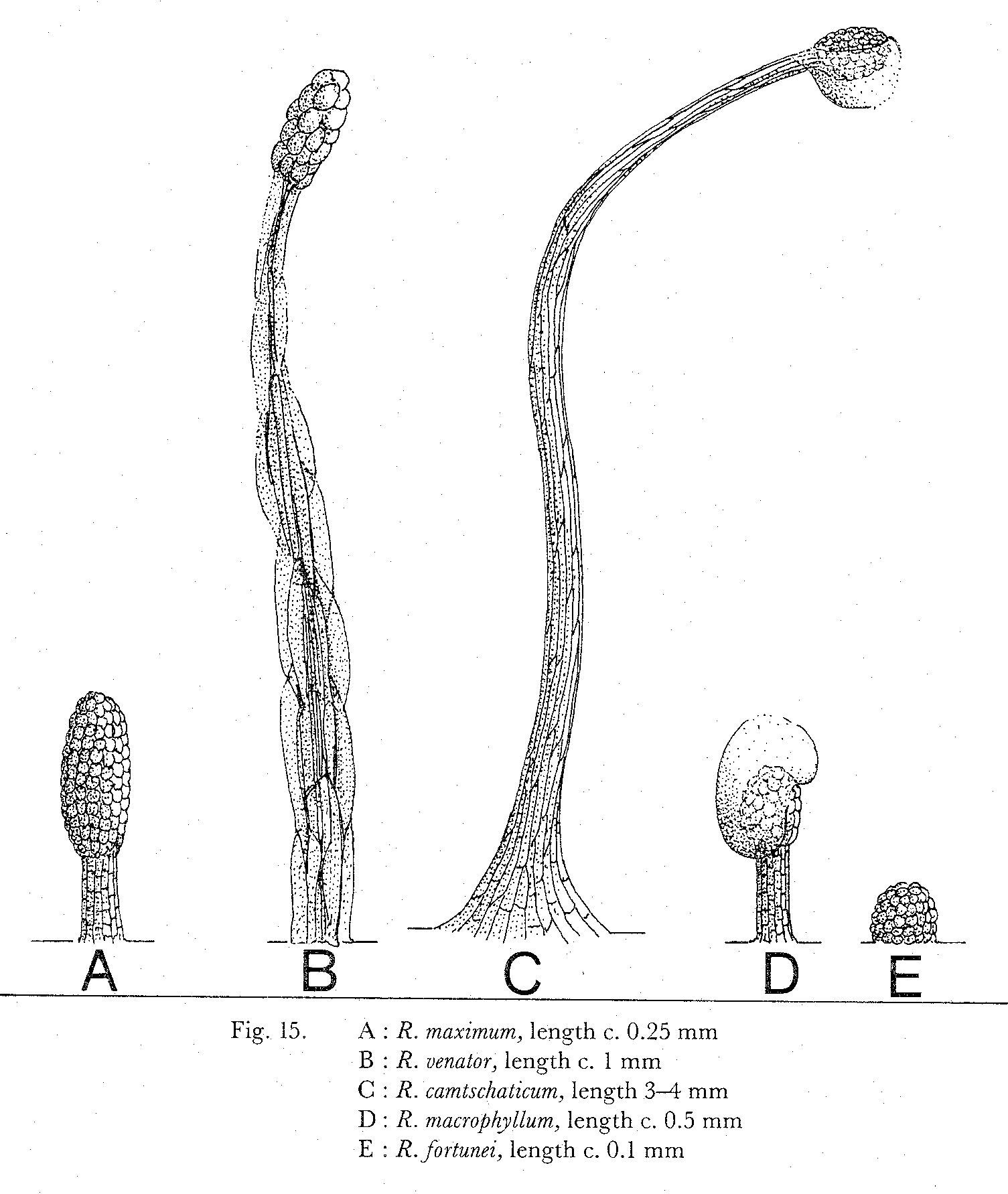
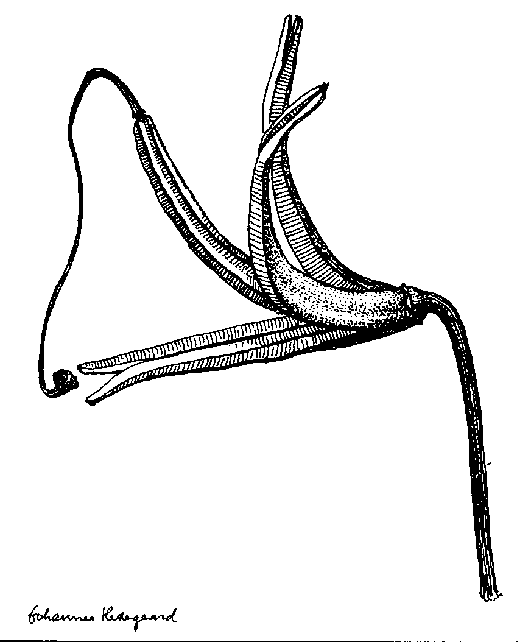
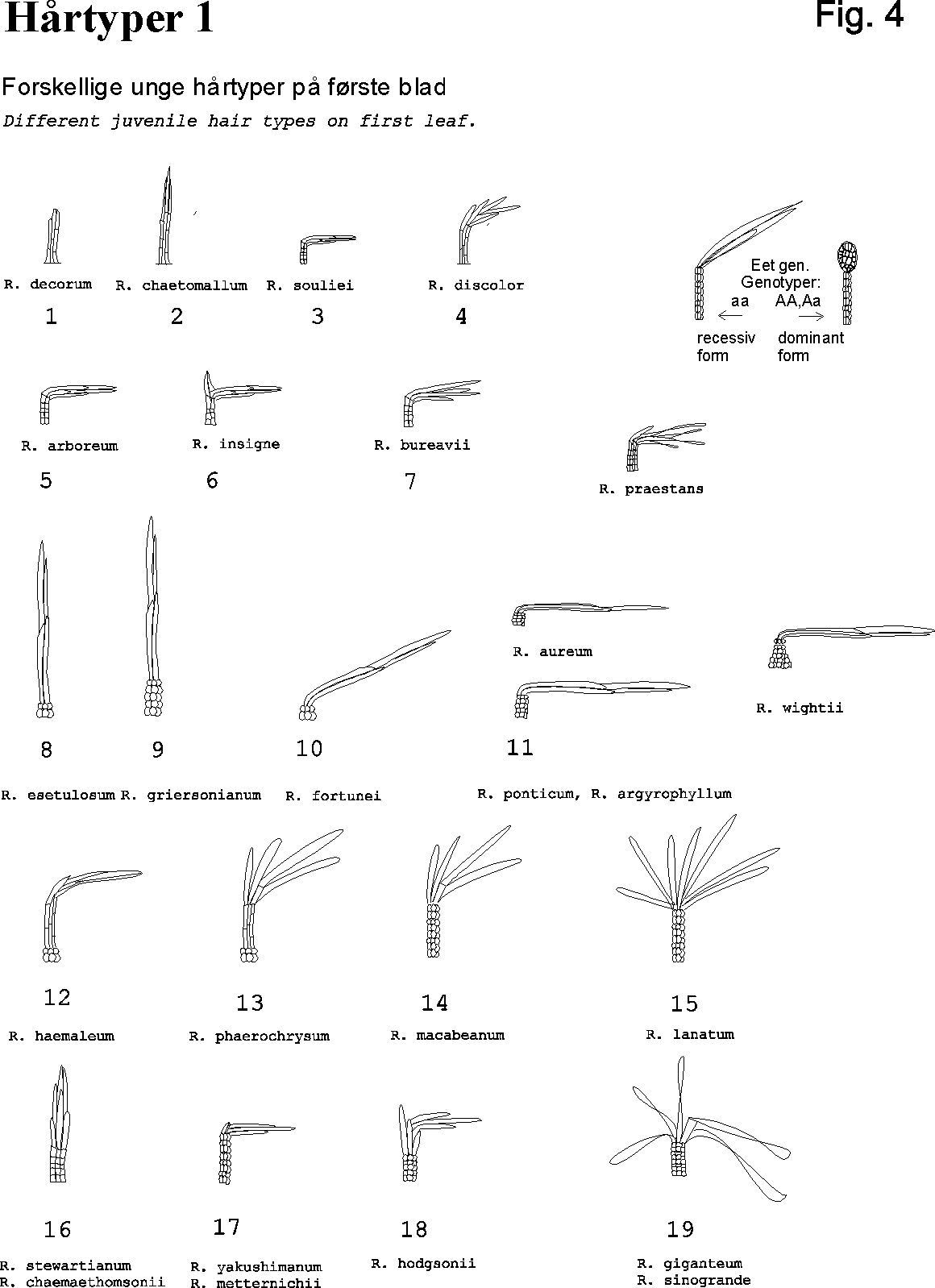
- Different eksamples og a glandular hair
- Eksample of different capsules
- Examples of different capsules
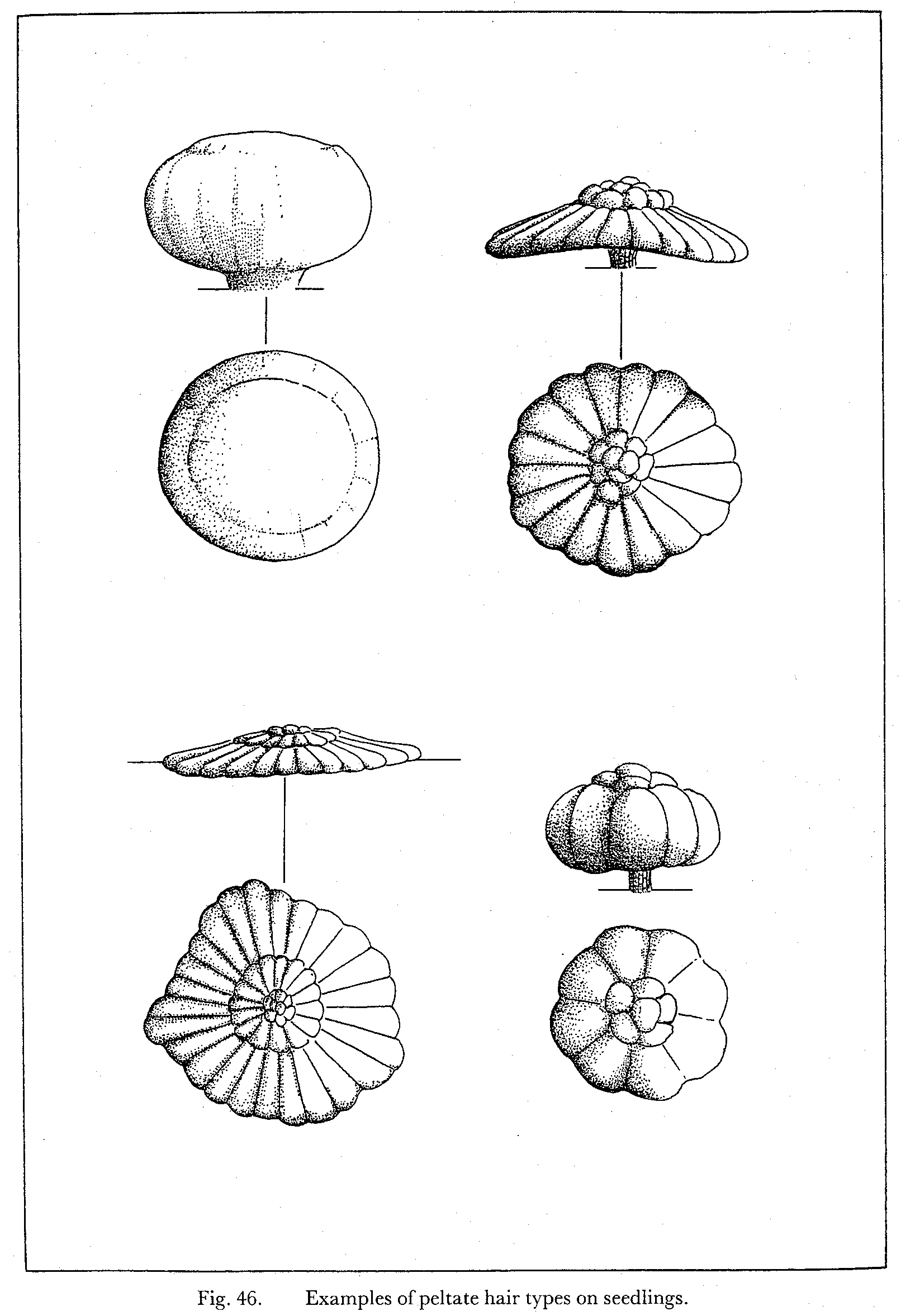
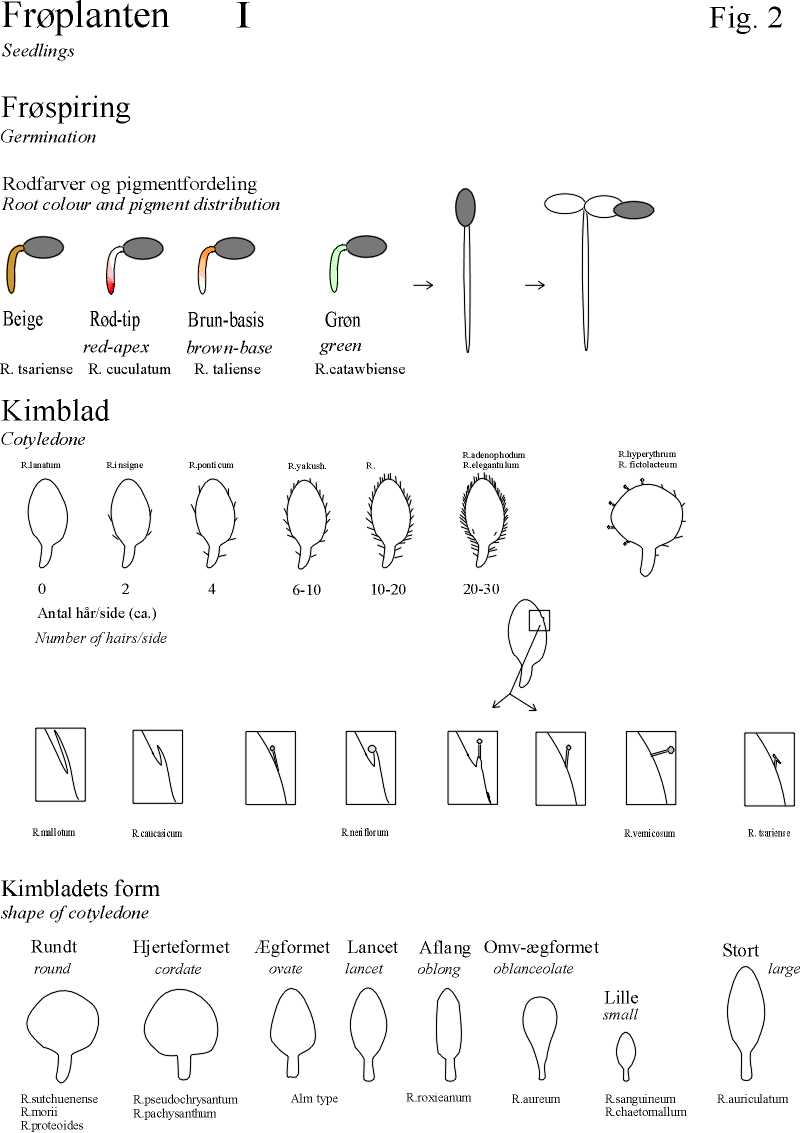
- Eksample of scales on seedlings
- Open dry capsule
- Cut of a capsule
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
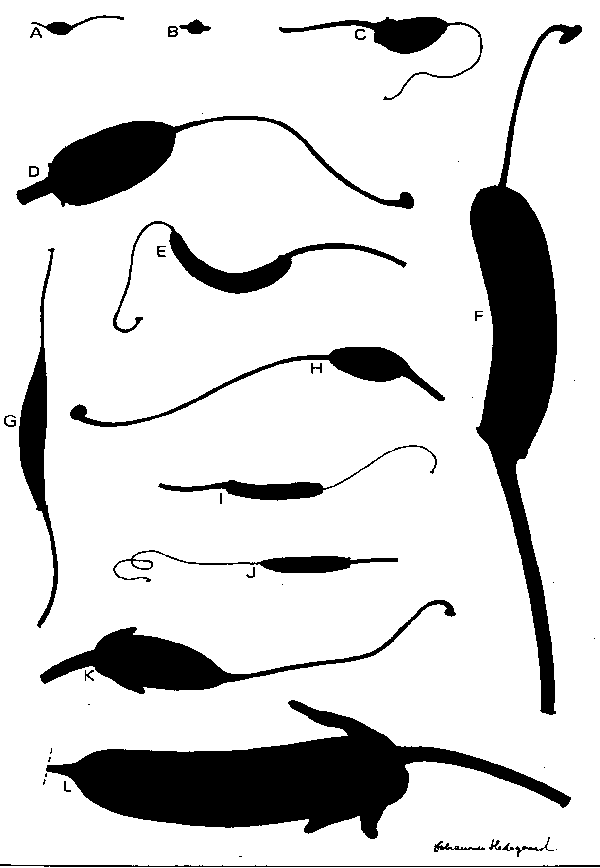{kind=link}
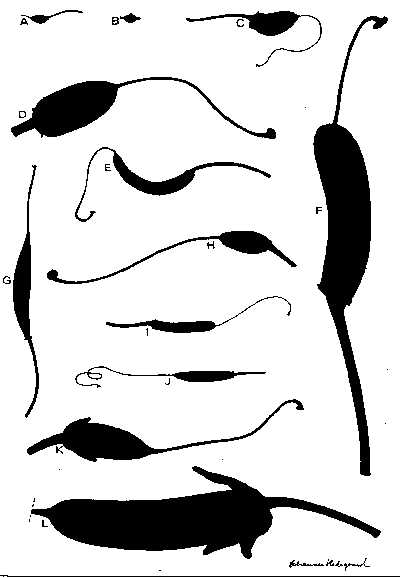{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nærbilleder af detaljer, Klaus Yde
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
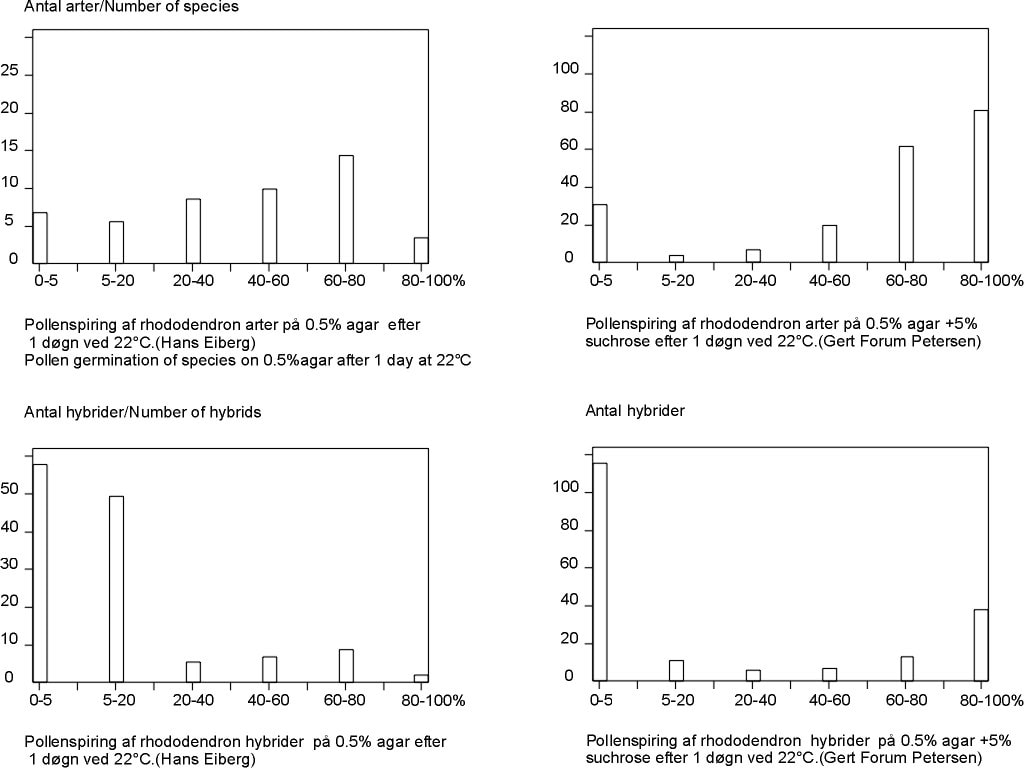
- Pollenspiringsforsøg på agar
- Pollenspiring på vand-agar: Resultater på hybrider. Polyploidy studies
{kind=link}
HVAD ER EN ART?
af Hans Eiberg
Ved møder i Rhododendronforeningen er der tit snakket om, om denne eller hin plante er en hybrid eller en art. Med den nye nomenklatur af D. F. Chamberlain er artsantallet blevet skåret ned, nye navne indført og arter ført fra den ene gruppe til den anden. I H. Davidians bog er nævnt ca 1000 arter, Chamberlain nævner 700, medens K. Cox giver udtryk for, at der måske kun er 500 arter indenfor rhododendronslægten. Hvorfor er det så svært af finde ud af, om en plante er en art eller en hybrid?
En af årsagerne hertil kan nok tilskrives den konkurrence, der blev skabt fra 1900 -1930 da indsamlerne E. H. Wilson, G. Forrest, F. Kingdon-Ward, J. F. Rock og R. E. Cooper kappedes om at introducere nye arter. Det lykkedes for G. Forrest fra 1904 – 1930 under 7 ekspeditioner at beskrive ca. 250 arter og hver ny indsamlet art blev honoreret med 400 pund? Nye forsøg på at genfinde disse dengang hastigt indsamlede arter / hybrider har ved de seneste indsamlinger i samme områder vist sig at være vanskeligere end ventet. Udbredelsen af mange arter, som førhen var sjældne, er nu yderligere begrænsede f.eks for clementinae og proteoides vedkommende. Ofte er biotoperne ændret eller ligefrem opdyrkede.
Davidian’s bøger
I Davidian’s bøger er arterne beskrevet ud fra herbariematerialer indsamlet af de ovenfor omtalte pionerer og planter fra plantesamlinger, især i England, hvor planter fra pionerernes indsamlede frø gror. Der kan sættes spørgsmålstegn ved mange af disse arters ægthed, som også Davidian var usikker på, såsom erythrocalyx og glaucopeplum. Disse “arter” menes nu at være hybrider. For erythrocalyx vedkommende vokser denne som enkelte planter i store populationer af wardii og selense, medens glaucopeplum vokser mellem phaeochrysum og aganniphum. Også flavorufum er nævnt som voksende mellem arterne phaeochrysum og aganniphum, medens den i andre områder vokser i store ensartede populationer. En af årsagerne til denne tvivl om artsantallet, er nok afstedkommet af, at artsdefinitionen er vag og omstændelig og er blevet ændret med tiden. Vi vil herunder beskrive lidt af artsbegrebets historiske udvikling.
HISTORISK UDVIKLING
Carl von Linné (1707-78), grundlægger af den videnskabelige botanik og systematik, opdelte systematisk forskellige planter (og dyr) i arter, samlet i slægter, der igen samles til familier, ordener, klasser og rækker. Hans plantesystem (seksualsystem, byggende på antal støvdragere mm) var velegnet som nøgle til bestemmelse af blomsterplanter. Planterne fik et dobbeltnavn, sammensat af et slægtsnavn og et artsbetegnelse f.eks R. ponticum.
Charles Darwin
Charles Darwin publicerede i sin bog, Arternes oprindelse, i 1859, at organismer ikke reproducerer sig identisk, men i forskellige variationer, af hvilke mange er arvelige – at kun de organismer overlever, som er i stand til at tilpasse sig de naturlige omgivelser – at alle organismer derfor fører en kamp for tilværelsen på grund af det store fødselsoverskud – at kun de bedst egnede overlever denne kamp – de livsformer, vi kender idag, har udviklet sig fra en eller få simple urformer (stamtræet der forgrener sig) – geografisk isolation var essentiel for artsdannelsen. Dvs. arterne vi kender idag er ikke stabile former, idet de stedse må tilpasse sig nye forandringer i omgivelserne, herved skabes en evolution (udvikling).
Omgivelserne, levevilkårene har ændret sig i tidernes løb og det har bevirket, at former for liv, som klarede sig udmærket i een tidsperiode ikke kunne overleve en anden på grund af klimaændringer; medens andre, som tidligere havde haft vanskeligt ved at klare sig, nu fik levevilkår, som passede dem bedre, og de levede så videre. Hverken dyr eller planter kan i nævneværdig grad “tilpasse sig” omgivelserne – i den forstand, at de kan udvikle de nye egenskaber, som de har brug for, men naturen udvælger de individer, der besidder de egenskaber, der er behov for. Darwins teorier er endnu idag grundlaget for videnskaben om arternes opståen og udvikling.
J. H. Wanscher
I 1942 i J. H. Wanscher’s bog: Arvelighedsordbog, beskrives en art som (citat):
“Art er et biologisk enhedsbegreb omfattende de individer, der har en sådan grad af lighed i de væsentlige egenskaber (blomster bygning o.s.v), at det har syntes naturligt at give dem samme navn. Artsadskillelsen er ofte noget vag og kan derfor diskuteres. Det er naturligt at antage, at de individer, der udgør en art, har samme oprindelse og har udskilt sig som en art fra tidligere eksisterende. Individer tilhørende samme art er så godt som aldrig ens hverken genotypisk (f.eks. to planter af samme art der ser ens ud, kan godt være forskellige, idet de kan indeholde flere eller færre vigende gener) eller fænotypisk (to planter af samme art, kan godt se forskellig ud).
Disse varianter lader sig ofte gruppere i subspecies, underarter, der i visse tilfælde adskiller sig så øjensynligt fra arten, at de betragtes som selvstændige arter. Ud fra denne betragtningsmåde, at arterne i naturen kun repræsentere former, der har hævdet sig overfor den naturlige selektion, har Turesson (1922) foreslået at kalde de naturstabiliserende arter (Linnéiske arter) for økospecies, som igen overfor de varierende voksekår har reageret ved at opdele sig i en række arveligt fikserede enheder, økotyper, der er i overensstemmelse med kårene. Med betegnelsen Coenospecies betegner han artens fulde variationsområde;..”
Artsbegrebet
Senere 1967 er artsbegrebet præciseret af I. Linde-Laursen og J. H. Wanscher med tilføjelsen: En gruppe individer af fælles oprindelse med en i hver generation tilbagevendende, genkendelig hovedtype eller sum af karaktertræk. En sådan konkret art vil, når den har bredt sig over større områder med vekslende vilkår og med fra hinanden isolerede populationer, komme til at fremtræde som en række til de forskellige underområders forhold særligt tilpassede former, racer eller økotyper, der skønt indbyrdes skelnelige dog er indbyrdes frugtbare f.eks. R. reticulatum og dens mange varianter i Japan. Modsætningsvis vil krydsninger med individer af forskellige arter være vanskelig, således at det sterile eller manglende afkom danner en beskyttende “artsbarriere”.
Den ovenfor givne artsdefinition er konkret, da den henviser til individer og deres afkom i mange led, det vi kalder frøstabilitet. Men søger man “arten selv” eller den “oprindelige art”, møder man uløselige vanskeligheder, thi arten vil da vise sig at være en fiktion, ingen plante vil være arten “i egen person”, men blot et individ, der rent systematisk henføres til den plads i det beskrivende system, som der henvises til ved hjælp af artsnavnet. Systemet i alle dets led er abstrakt – en samling beskrivelser for artens vedkommende knyttet til typeeksemplarer. Brugelige betegnelser for den konkrete art er øcospecies og coenopopulation, medens ordet art bør forbeholdes den abstrakte art.
SÅDAN! Ikke så underligt, at det er et svært spørgsmål at besvare!
Det har været nødvendigt at indføre genetik (betragte flere generationer) for at præcisere artsbegrebet fra 1942 til 1965. I nyere tid har genetikkens forskningsresultater haft stadig større og større betydning for artsbegrebet, idet det har været muligt at se enkeltstrukturer i kromosomernes opbygning mellem de forskellige dyre arter og mange plantearter. Da rhododendron kromosomer er meget små, har det været vanskeligt at se forskelle i strukturen, men antallet er ens for de fleste arters vedkommende (oftest 26=2N eller 52 stk=4N).
Hans Eibergs opfattelse
Min (HE) egen “genetiske” opfattelse af ægte etablerede arter er:
En samling individer, hvor generne for de forskellige egenskaber ligger i samme rækkefølge på hvert eneste kromosom. (altså med ens genkort). Der er ingen fysiske kromosomale barrierer, når genkortene er ens. Hvis en plante har dannet nogle ekstra gener, der koder for nye egenskaber, som har betydning for dens overlevelse, må den tilhøre en ny art. Varianter er planter, der har har samme genkort som arten, men et eller flere gener har en ny variation.
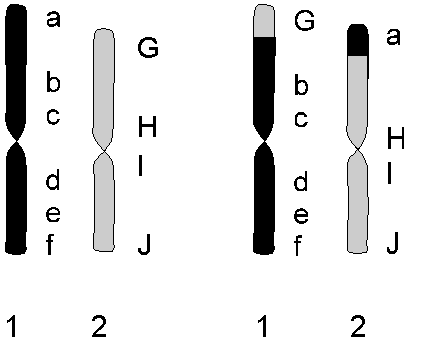
mht. 2 forskellige kromosomer
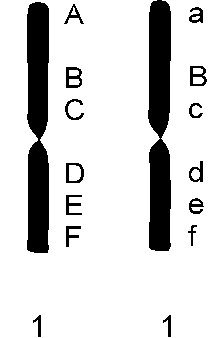
mht. 2 forskellige kromosomer
A, B, C, D, E of F er gener der styrer forskellige egenskaber (f.eks. højde, bredde, længde, indumenttype, farve osv). A1 og A2 er forskellige, men styrer samme egenskab (f.eks mere eller mindre gul blomsterfarve).
Artsdannelsens mekanismer
Der kan tænkes mange årsager til at to livsformer skiller sig fra hinanden. Artsdannelsen forløber som en kombination af flere forskellige hændelser ofte efter samme rækkefølge f.eks.
- Forøgelse af populationen, overbefolkning (samtidig sker en forøgelse af de “mere eller mindre gode” mutationer, hvilket bevirker at den genetiske variation øges)
- Fødeknaphed medfører dødsfald og isolation af individer. Kamp for at overleve.
- Spredning (migration) af individer (dyr).
- Enkelte individer har held til at forplante sig mere end andre (selektion, tilpasning) på grund af udseendet, styrke, adfærd, lugte mm. (den som bedst tilordner sig miljøet overlever).
Det er en fordel, at den biologiske variation er så stor som muligt, især når omgivelserne ændre sig. Nogle individer med specielle egenskaber har nu større chancer for at overleve, og de overlevende, som gruppe er anderledes end den oprindelige gruppe, hvilket er et skridt på vej til dannelsen af en ny art.
Artsstabilisering indenfor dyreriget
Inden for dyreverden er kromosomkort ved at blive konstrueret for mange arters vedkommende. Alle menneskeracer har samme genkort, medens chimpanser, gorillaer mm har et lidt anderledes kort. Forskellen mellem de tæt beslægtede dyrearter er en eller få nykonstruktioner i genkortet. To kromosomdele kan f.eks have byttet plads. Ombytninger bevirker, at hybrider/bastarder mellem arter er mere eller mindre sterile, idet kromosomparring mellem uens kromosomer er stærkt begrænset. Der er beskrevet mange kromosomafvigelser hos mennesket, som giver sygdom. Men også raske personer kan have et andet genkort, men personerne har stærkt nedsat frugtbarhed og kan kun få børn med personer med samme genkort.
Artsstabilisering inden for planter
Planter kan jo ikke selv vælge deres partner, og derfor må de for at kunne overleve have specielle mekanismer for at kunne vokse, formere, isolere og beskytte sig imod fremmede planter af nærtstående arter.
Der kan tænkes på manglende pollenspiring af fremmed pollen på støvfanget og forskellig blomstringstid. Eller overleve ved at besidde gener, der giver resistens mod sygdomme (beskyttelse mod parasitter), speciel vækstregulering, frostresistens, næring, vandoptagelse mm.
Der er ingen endnu, der har lavet genkort over en eneste rhododendron art, men der er små forskelle i genkortene mellem nogle planter, så ægte arter er en realitet. For de stedsegrønne elepidote arter (Hymenanthes) gælder, at alle de beskrevne arter kan danne primær hybrider (ingen pollenspirings barrierer). Hybriderne har ofte nedsat fertilitet, da de danner færre frø og mindre pollen, der ofte har en meget ringe vitalitet og spireevne, hvorfor mange hybrider i naturen ikke er stand til at etablere og opretholde en selvstændig population.
Arterne holder sig konstante, især på grund af isolation (vokser på hver sin side af en bjergkam mm.), forskellig blomstringstid, forskellig frost og sygdomsresistens, samt at arterne sætter flere fertile frø end hybriderne. Biologer mener at det er logisk, at arter isolerer sig fra hinanden. Organismer, der er tilpasset en speciel livsstil, vil ikke have nogen fordel af at blande deres gener
med andre, som har udvikler sig i en helt anden retning. Derfor har en række forskellige isolerede mekanismer udviklet sig, så parring fortrinsvis kun sker mellem artsfæller.
VÆSENTLIGE EGENSKABER
De væsentlige egenskaber arter opdeles efter er nødvendigvis ikke de egenskaber, der er knyttet til arternes overlevelse i miljøet. Arter opdeles efter deres udviklingsmæssige baggrund. Dvs nærtstående arter har nærtstående genkort, er tættere i familie med hinanden, idet de har nærtbeslægtede forfædre (samme oprindelse).
- Overlevelse
Indenfor artsbegrebet tales om, at individer indenfor samme art har de samme væsentlige egenskaber. Det vil da være rimeligt at tænke på egenskaber, som har betydning for artens/planternes tilpasning til miljøet for, at de kan overleve. Her tænker vi på miljøfaktorer som lys (intensitet og dagslængde), temperaturvariationer gennem året, vindforhold, næringsforhold samt resistens mod skadedyr og svampe. De genetiske faktorer (artsegenskaber) skal derfor tilpasses disse mange forskellige ydre forhold, således at der for hvert miljøkompleks er tilpasset en bestemt art. Planter kan se næsten ens ud, hvis de er tilpassede samme miljø igennem årtusinder, men alligevel være langt fra hinanden mht slægtsskab (sukkulenter kan tilhøre forskellige plantefamilier). - Klassifikation
For Rhododendron er det især egenskaber som: blomstens opbygning, bægerbladets udseende, besiddelse af skæl, blomsternes placering på grenen, løvfældende blade, indumentets opbygning mm. De væsentlige egenskaber er egenskaber, der styres fra
mange gener i samarbejde med hinanden. Egenskaber der styres fra eet gen er uanvendelige som middel til definition af en art. Reguleringsgener (bestemmer længde, bredde, antal og styrke) er ikke gode til brug for arts definitioner.
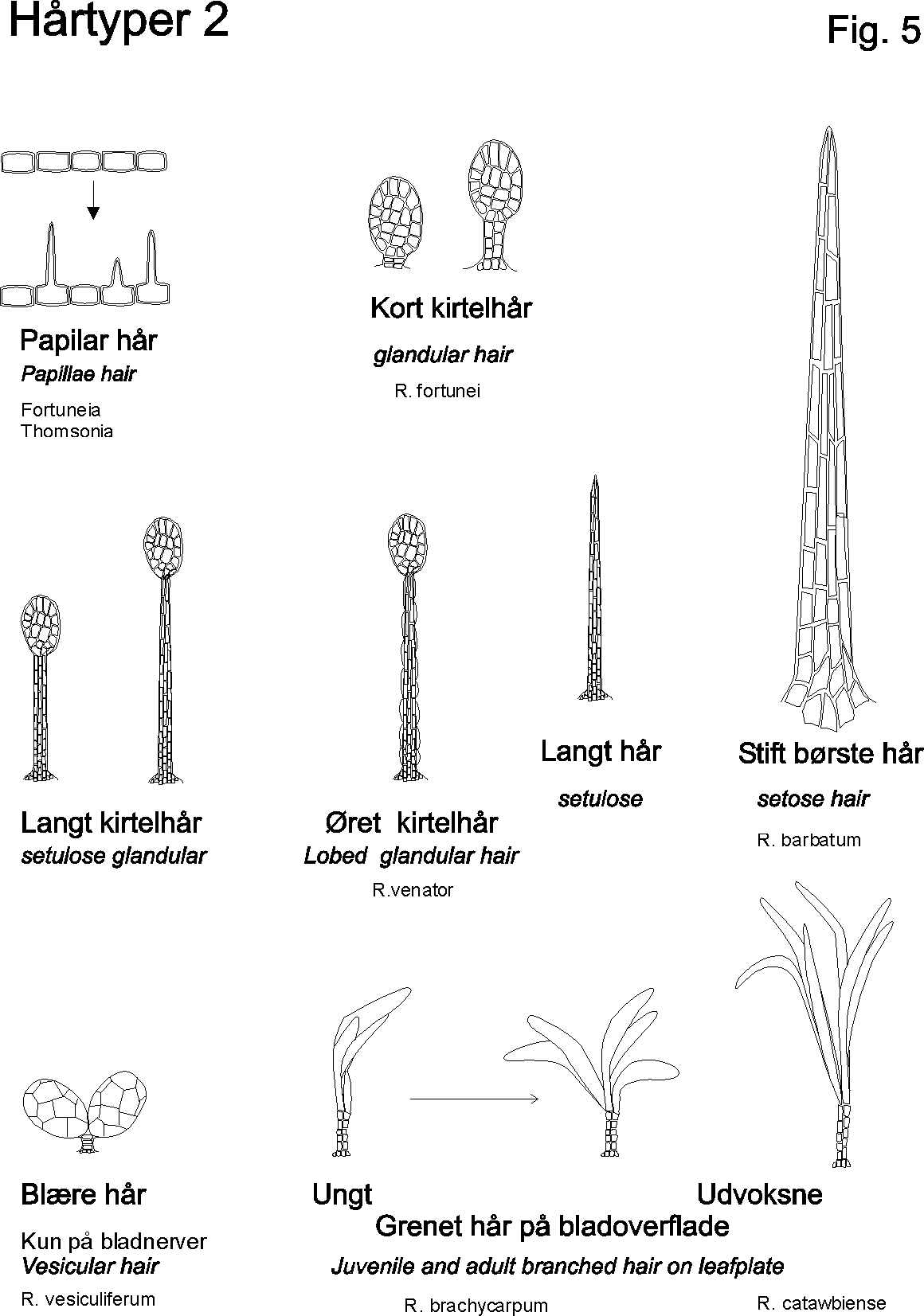
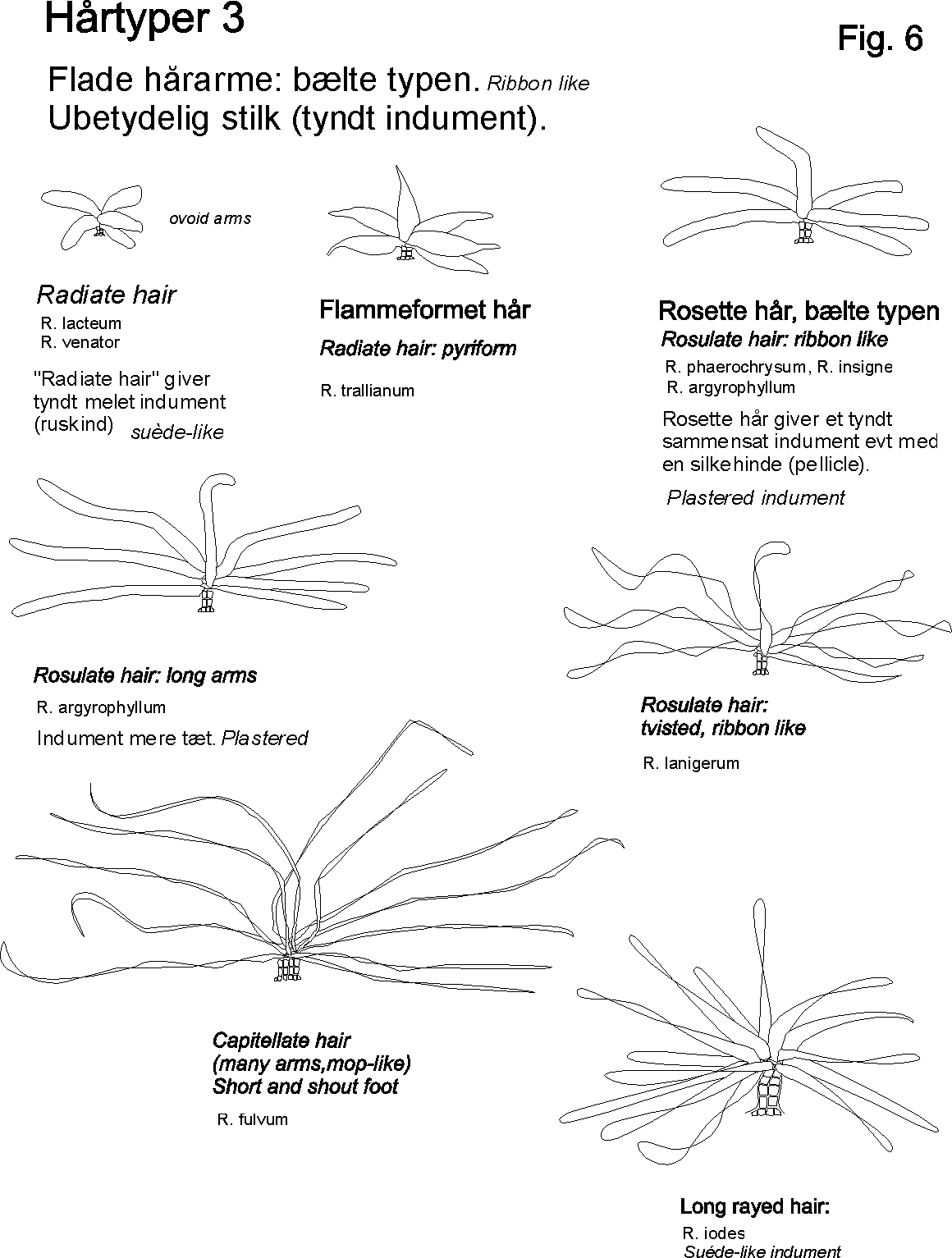
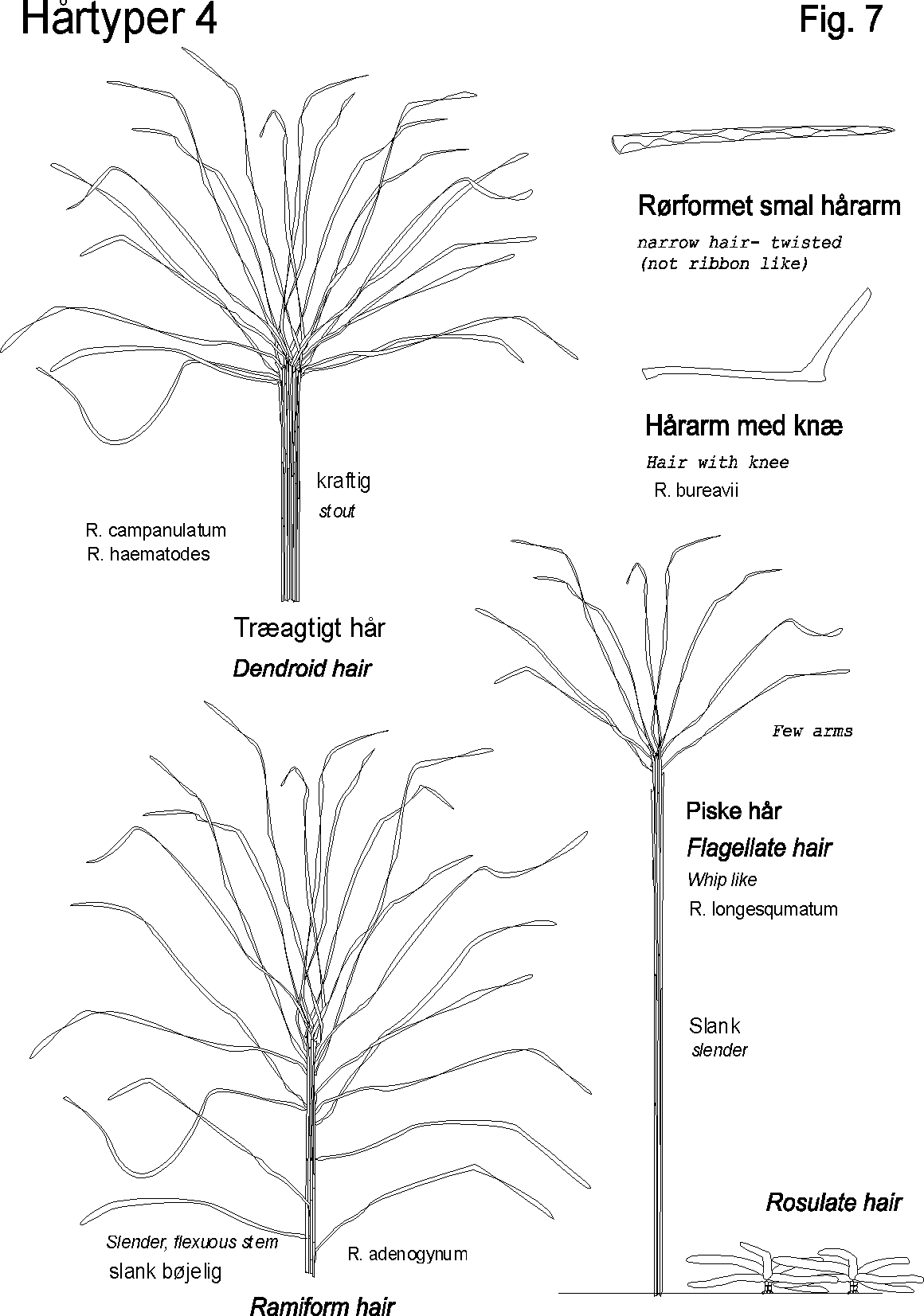
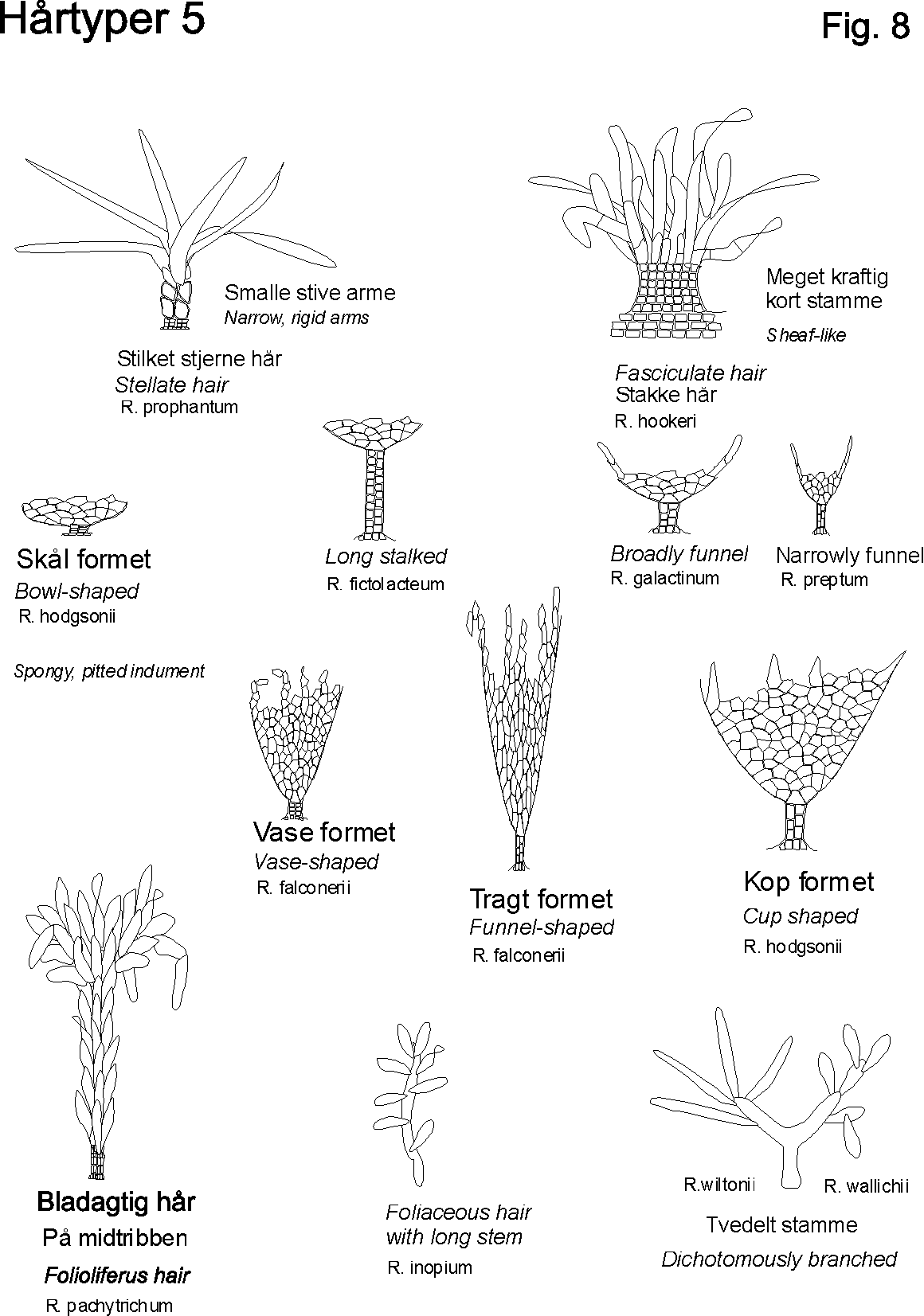
Egenskaber som ekstra / nye strukturgener, som andre planter ikke har er oplagte indikatorer til brug i artsbestemmelsesnøgler. F.eks kan indenfor håropbygningen nævnes dækhår(skæl), stjerne-, mop-, kirtel-, rør-, grenet-, børster-mm.
Man kan godt betragte de nu beskrevne arter som selvstændige arter, hvis hver art er i besiddelse af nye gener eller genkombinationer, som adskiller sig fra de øvrige arter og især, hvis de har betydning for den respektive arts overlevelse.
Truede arter
De planter f.eks clementinae, der kun kendes fra enkelte indsamlinger, er problematiske mht artsbegrebet. Kan een / to planter kaldes for en art? Ikke efter definitionen. Nu har vi jo opformeret den fra frø (selvbestøvning?) og samtidig kan der være sket nymutationer i det opformerede materiale. Derfor må den efter vores mening betegnes som en art, hvis frø- afkommet ikke afviger væsentligt fra forældre/forældrene. Ved denne opformering, har vi måske reddet en tidligere art i naturen fra at uddø for menneskeheden, ved at holde den i live i vores egne haver.
Hvordan kan vi bestemme om en plante er en hybrid eller en art
Når vi står overfor en plante og skal bestemme hvilken art den tilhører, bruger vi først vores kendskab til plantefamilien og kan i nogle tilfælde straks se hvor planten hører hjemme alt efter vores erfaring. Ellers må man bruge en plantebestemmelsesnøgle eller fotografier / tegninger. De bestemte karakteristika planten har bør altid sammenlignes med artsbeskrivelsen efter en af H. Davidians bøger I-III, The Rhododendron handbook, P. A. Cox: The Larger species of Rhododendron, Dwarf Rhododendrons, F. C. Galle: ” Azalea”.
Planten kan sammenlignes med andre planter, der er bestemt som hørende til arten. Hvis der ikke er overensstemmelse mellem de væsentligste træk, må der straks tænkes på, at planten kan være en hybrid. Der kan undersøges om planten normalt sætter rigeligt med frø, hvis den er fremmedbestøvet. Om der opnås et ensartet afkom (tager flere år). Om plantens pollen er spiringsdygtigt (helst >50% spiringsevne). Desværre har vi ikke DNA test, der kan bestemme art/hybrid sammenhængen, men det kommer. Det er endnu kun muligt kemisk at bestemme indholdet af forskellige flavonoider, vokstyper mm i planten for at afsløre, om det er en hybrid (Späetmann, B. A. Knights). Hybrider vil være mellemformer, når de findes i naturen og næsten altid vokse sammen med forældrene i områder, hvor krydsbestøvning forekommer (bier / biller og fugle må være tilstede).
Arvelige træk kan også være en god hjælp. Fx er mange arter beskrevet som hybrider nogle steder i litteraturen fx. R. wongii. Men arvegange viser at det næppe er sandsynligt, idet R. wongii har hår på griflen (recessivt) og en af de foreslåede forældre (R. ambiguum) har ikke disse hår. Nogle vil så påstå at det er en F2 hybrid der er stamfaren til R. wongii, men hybrider plejer at uddø i naturen i skarp konkurrence med arterne der sætter flere frø. Også dominante egenskaber kan vise om en plante er en hybrid. fx hår på ovarie/støvdragere er dominant arvelige.
RESUMÈ
Rhododendronfamilien er opdelt i forskellige underfamilier, slægter, arter, underarter, og varianter ud fra en sammenligning og optælling af fænotypiske træk, og muligheder for hybridisering. Denne fordeling har i de sidste 100 år været genstand for utallige diskussioner mellem botanikere verden over.
Efter at have beskrevet definitionerne på artsbegrebet er vi enige i, at indenfor Hymenanthes underfamilie (elepidote stedsegrønne planter, f.eks yakushimanum) er der et nært slægtsskab mellem de beskrevne arter, idet alle? / de fleste arter kan danne hybrider med hinanden. Der er så nært slægtskab, at hybriderne er mere eller mindre fertile! dvs at der kan sættes spørgsmålstegn ved, om der er mere end een “rigtig” art i denne gruppe, hvis man bruger fertilitetskravet på hybriderne som artskriterium.
Da plantegruppen ikke er så gammel (mellem 165-200 millioner år (P. Cox. The larger species of Rhododendron..) har artsdannelsen ikke nået at stabiliseret sig ved at danne effektive barrierer (forskelle i kromosomernes genkort). Skal vi derfor kalde alle arter indenfor Hymenantes for underarter? og bruge artsnavnet som underartsnavn, fordi alle hybriderne er fertile? Det vil være ærgerligt og urimeligt. Hvis f.eks forestii repens skal tilhøre samme art som sinogrande. Vi må derfor finde os i, at færre forskelle end normalt (og accepterer næsten ens genetiske kort mellem arterne) anvendes som begrundelse for at opdele to nærtstående planter i to forskellige arter.
Hans Eiberg
og Gert Forum Petersen (korrektur)
————- senere noter i 2002 ——————-
One of the best known species defintion is Ernst Mayr’s biological species concept. In short:
“A species is a group of interbreeding natural populations that is reproductive isolated from other such groups”
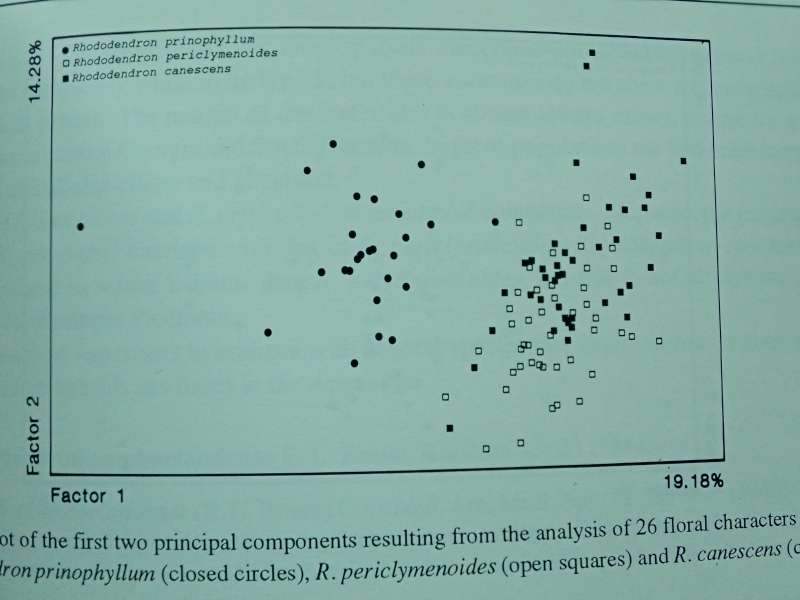
GENETIK
- Arvelighed af hår på griflen, ovarie og støvdragere
- Introduktion i artsbegreb 2009 , se ovenfor
- Mendel’s paper in 1865 – 95% af alle morfologiske træk nedarves kvantitativt (polygenet) ikke Mendelsk
- How many genes are involved for a trait? Hans Eiberg (PDF), Engelsk
- Genetiske regler og deres anvendelse (Mendelske)
- Pollenspiring. Arter og hybrider sammenlignes på vand og sukker
- “Moder-effekten” har betydning for vitaliteten – Imprintning. Germanica x R. fortunei (TV). R. fortunei x Germania (TH) Foto: Mogens Rasmussen
- Hvordan finder man et gen for en egenskab vha kobling
{kind=link}
TAXONOMI
Nomenklatur
Evolution
- About evolution – Wikipedia
- The calendar of the Earth- Steno
- Evolutions theorien (Danish) – wikipedia
Klassifikation – systematisering af rhododendronslægten (bl.a. med DNA- analyser)
af Hans Eiberg, Universitetet i København.
(abstract til Nordisk rhododendron symposium, Bergen juni 2003)
Under de mange ekspeditioner i Asien fra 1900-1940 blev der indsamlet store plantematerialer. Dette blev overdraget til Edinburgh Botaniske Have til videre undersøgelse og karakterisering under ledelse af prof. Balfour. Udfra de kendetegn rhododendronplanterne besad blev disse anbragt i 27 foreløbige grupper (Serier). Serier, fordi planterne var vanskelige at sætte i det gældende system på grund af manglende og få data mht. udbredelse og viden om hvilke træk der var væsentlige til dette brug. De træk, som blev anvendt var ustabile træk, som i mange tilfælde er fordelagtige for planten her og nu, for at denne kunne overleve i naturen.
Det første system, der kun anvendte den vedtagne nomenklatur blev foreslået af Sleumer (1949). De kendetegn han anvendte til systematisering af slægten er de samme, som er anvendt af senere forskere, men med evt. en lidt anden vægtning. Et forsøg med andre markører (farvestofenzymer) blev foreslået af Spätman, men hans system stred mod nomenklaturen, idet Serie betegnelsen også blev anvendt.
Botanikernes fornemste resultat er foruden en beskrivelse af arterne, også en udfærdigelse af en anvendelig nøgle til bestemmelse af de forskellige arter i slægten. Det er dog stadig gældende, at planter der findes i naturen ikke altid kan bestemmes efter disse udfærdigede nøgler, hvor de klassiske træk anvendes. I dag kan man bestemme rækkefølgen af baserne i DNA (sekventering) og dermed påvise miljøneutrale variationer til brug for systematisering af slægten. Denne metode kan på en gang påvise snesevis af væsentlige variationer. Dette skaber grundlag for at indordne planter / arter i et system der følger evolutionen. Metoden er ideel til at opbygge et sandt system, men upraktisk at anvende til plantebestemmelser i marken.
Når flere DNA områder er undersøgt vil mængden af data kunne vise den bedste relation af slægtskabet til brug for en gruppering af planter i det gældende system: subgenus, sektions, subsektions og endvidere hvilke planter der er hybrider. En samlet DNA profil kan løse en del af problemerne med at definere arter der er tæt relaterede (alliancer). For at udføre alt dette arbejde må fejlkilder overvindes og så mange arter og DNA stykker undersøges som muligt.
DNA-analyser, Phylogenetic
Phylogenetic trees published in “Rhododendron i Danmark i 25 år; 1999 by Hans Eiberg” 4/5-1998 Dansk version (2.3Mb)
Phylogenetic trees published in “Rhododendron i Danmark i 25 år; 1999 by Hans Eiberg” 4/5-1998 Engelsk version (0.3Mb)
Analysis of 20 ITS sequencies by hand. Hans Eiberg in 1997
Analysis of 28 ITS sequencies by DNAML fig 2 modificeret
Analysis of ITS Penthanthera sequencies by DNAPARS – fig 5
Analysis of 29 ITS sequencies in 1998. DNAML modificeret
Analysis of 20 MatK sequencies by DNACOM.
{kind=link}
{kind=link}
{kind=link}
Hvordan laver man et stamtræ udfra DNA sekvenser
NCBI Taxonomi: DNA sequencies
- Rhododendron familiens stamtræ
- BioBank DNA
- 29 ordnede DNA-sekvenser udfra 29 arter
- Fylogenetiske træer (mange) udfra rhododendron arter (ITS området ved 5.8rRNA genet) (972K)
- Fylogenetisk træ lavet udfra 10 rhododendron arter (matK genet) – 14/1-1998
- Fylogenetiske træ lavet udfra udfra 31 rhododendron arter (matK gene) – 4/5-1998
- Find arten som DNA sekvensen stammer fra – og se stamtræet
- About phylogenetics
- Glossary about phylogenetics
- Download programs for phylogenetics, CMAP, COMPARE, PHYLIP
Some scientific articles
- Google search
- Spatiotemporal Evolution of the Global Species Diversity of Rhododendron 2021. Stort stamtræ
- Goetsch et al. 2005 abstract
- matK genet – Molecular systematics of Rhododendron subgenus Tsutsusi
- A decade of progress in plant mol phylogenetic